

Artículos originales
Crecimiento foliar y comportamiento diario de la fotosíntesis en plantas de fique cv Ceniza (Furcraea cabuya Trelease)
Foliar growth and daily behavior of net photosynthesis in fique plants cv Ceniza (Furcraea cabuya Trelease)
 Jorge Cadena-Torres jcadena@agrosavia.co
Eduardo Barragan Quijano
Jorge Luis Romero Ferrer
Kelly Mercado
Jorge Cadena-Torres jcadena@agrosavia.co
Eduardo Barragan Quijano
Jorge Luis Romero Ferrer
Kelly Mercado
Temas Agrarios
Universidad de Córdoba, Colombia
ISSN: 0122-7610
ISSN-e: 2389-9182
Periodicidad: Semestral
vol. 26, núm. 2, 2021
Recepción: 21 Junio 2021
Aprobación: 14 Septiembre 2021

Resumen: El presente estudio se llevó a cabo con el objeto de monitorear el crecimiento foliar, la emisión de hojas y el comportamiento diario de la fotosíntesis en plantas de fique cv Ceniza. Para lo anterior, en un cultivo comercial se seleccionaron plantas de fique a las cuales se les realizó un monitoreo semanal para determinar las ganancias en número, longitud, ancho y grosor de las hojas, durante un período de 154 días. En dos oportunidades durante el periodo experimental, se realizó también un monitoreo cada hora a las tasas de intercambio gaseoso durante los periodos de luz y oscuridad. Los resultados de crecimiento permitieron estimar un filocrono promedio de 8,65 días. El monitoreo a las tasas de intercambio gaseoso indicaron una baja conductancia estomática durante el período de luz. Sin embargo, a partir de las 15:30 h, la conductancia estomática se incrementó sustancialmente, manteniéndose a niveles máximos durante todo el período de oscuridad, para posteriormente decrecer a partir de las 09:30 h, del siguiente periodo de luz. Se encontró una alta correlación de la conductancia estomática con el déficit de presión de vapor y la temperatura foliar. Las máximas tasas de fijación de CO. detectadas variaron entre 12,2 y 14,5 µmol m-2 s-1, las cuales se registraron entre las 19:19 y las 19:26 h, durante el período de oscuridad. Estos resultados corroboran el metabolismo CAM de las plantas de fique cv Ceniza.
Palabras clave: Crecimiento foliar, Fotosíntesis, Filocrono, In- tercambio gaseoso, Plantas CAM.
Abstract: The present study was carried out to monitor leaf growth, foliar emission, and the daily photosynthesis behavior of sisal plants cv Ceniza. In a commercial crop, sisal plants were selected and monitored weekly to determine increase in number, length, width, and thickness of the leaves, during a 154 day period. On two occasions during the experimental period, gas exchange rates were also monitored during the light and dark periods. The results allowed determining a phyllochron of 8.65 days. Gas exchange monitoring detected low stomatal conductance rates during the light period. However, starting at 15:30 h, stomatal conductance substantially increased, remaining at maximum levels throughout the dark period, and, subsequently, decreased from 09:30 h, the following light period. A high correlation of stomatal conductance with vapor pressure deficit and leaf temperature was found. Maximum CO. fixation rates varied between 12.2 and 14.5 µmol m-2 s-1, recorded between 19:19 and 19:26 h. The results corroborate the CAM metabolism of sisal plants cv Ceniza.
Keywords: Foliar growth, Photosynthesis, Phyllochron, Gas exchange rates, CAM plants.
INTRODUCCIÓN
El fique es una planta originaria de la América Tropical en donde se utiliza principalmente para la producción de fibras naturales para la fabricación de sacos, cuerdas y artesanías, pero también como planta ornamental, para cercas vivas y control de la erosión en laderas (Yu, 2005; Anandjiwala y Maya, 2010). En Colombia el uso de la fibra se ha concentrado en la elaboración de sacos para el empaque y comercialización de productos agropecuarios, principalmente café, cacao, papa y hortalizas (Casierra-Posada y González, 2009).
En Colombia las principales especies cultivadas pertenecen al género Furcraea, recientemente descrito y reagrupado por Eggli y Nyffeler (2020), en donde las especies más comunes son Uña de Águila (Furcraea macrophylla Baker), Negra Común (Furcraea gigantea Ventenat), Borde de Oro (Furcraea Castilla), Ceniza (Furcraea cabuya Trelease) y Sin Espínas (Furcraea andina Fritsch). El área cultivada se ubica principalmente en los departamentos de Nariño, Cauca, Santander, Antioquia y la Guajira, con un área de siembra registrada para el año 2018 de 18.982 hectáreas (Agronet, 2020).
Su cultivo es muy común en regiones con alta participación de minifundio, en donde la fibra producida aporta recursos importantes para la sostenibilidad económica de la familia. Aun cuando la tendencia indica un incremento en las áreas de siembra, en los años recientes se ha presentado un incremento en la floración espontánea de las plantas lo que ha ocasionado muerte y diminución de las plantaciones activas y en producción. Se considera que lo anterior puede estar siendo ocasionado por el calentamiento global.
Pocos estudios se han adelantado en Colombia acerca de la fisiología del crecimiento y desarrollo de la plata de fique, así como de su comportamiento fotosintético y su relación con la temperatura bajo condiciones de cultivo, con adecuada disponibilidad de agua y nutrientes (Casierra-Posada y Gómez, 2008; Casierra-posada y González, 2009). En estudios adelantados por Casierra-Posada y González (2009), se comprobó que las platas presentan ritmos circadianos en la acidez y los valores de pH de las hojas, que permiten inferir su comportamiento como planta con metabolismo CAM. De igual forma, los estudios de anatomía foliar adelantados por Murillo-Serna, JS et al., (2018), sugieren una estructura y arreglo celular típico de plantas con metabolismo CAM (Murillo-Serna, JS; Rincón-Baron, EJ y Alzate-Guarín, 2018). Con base en lo anterior se asume que las plantas de fique presentan alta conductancia estomática y asimilación del CO2 durante el periodo de oscuridad. Para corroborar lo anterior, la presente investigación se llevó a cabo con el objeto de estudiar el crecimiento foliar y comportamiento diario de la fotosíntesis de las plantas de fique cv Ceniza bajo condiciones de campo.
MATERIALES Y MÉTODOS
Localización
La presente investigación se realizó bajo condiciones de campo en el municipio de San Vicente Ferrerer, en el departamento de Antioquia, zona de tradición en la producción de fique en el departamento. El sitio donde se realizó la investigación se encuentra a una altitud de 2.350 msnm, con suelos de reacción ácida, con alto contenido de materia orgánica, alto contenido de fósforo, calcio, potasio y zinc; medio contenido de azufre, boro y cobre, y bajo de magnesio y manganeso. Durante el desarrollo del experimento las temperaturas nocturnas fluctuaron entre 9,8 y 15,8 °C, y las diurnas entre 19,8 y 25,2 °C, con una oscilación que varió entre 5,4 y 14,8 C.
La temperatura media registrada durante el período experimental fue de 17,8 °C, mientras que la precipitación acumulada fue de 1936.6 mm. Para los cálculos fisiológicos se desconoce la temperatura base del fique, por lo que las variables de evaluación se presentan usando el tiempo cronológico transcurrido desde el inicio de las mediciones.
Material vegetal
Las evaluaciones se realizaron en una plantación comercial de fique cv Ceniza (Furcraea cabuya Trelease). El cultivo utilizado para las mediciones presentaba variabilidad en la edad de las plantas, por lo que acorde al criterio de agricultor se conformaron grupos de plantas, de acuerdo a su edad a su edad (<10 años, 11-20 años y >20 años) y se realizaron monitoreos sobre los tres grupos de plantas, a manera de repeticiones. Las plantas de mayor edad presentaban registros de cosechas previas de hojas.
Variables evaluadas
Número de hojas emitidas: Se registró semanalmente el número de hojas presentes en las plantas durante un periodo experimental de 154 días y con los datos obtenidos se calculó el número de hojas emitidas, mediante la siguiente relación.
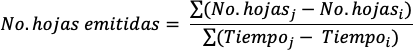
Donde i y j representan periodos sucesivos de muestreo.
Tasa de aparición foliar (TAF): se calculó como la pendiente de la función lineal que relaciona el número de hojas emitidas a través del tiempo cronológico transcurrido en el período experimental. Con la tasa sucesivas, mediante el inverso de la tasa de aparición foliar (Di Benedetto y Tognetti, 2016).
Crecimiento foliar:se dividieron las plantas en tercios (superior, medio y bajo), se marcaron las hojas de cada tercio y semanalmente se registró sobre la misma hoja, las ganancias en longitud, ancho y grosor de las hojas, por un período de 154 días. Las ganancias en longitud, ancho y grosor de las hojas se calcularon mediante la siguiente relación:
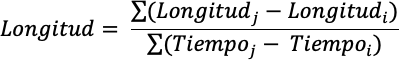
Donde i y j representan periodos sucesivos de muestreo.
Tasa de elongación foliar (TEF):la tasa de elongación foliar se calculó con base en las ganancias en longitud, ancho y grosor foliar en función del tiempo cronológico transcurrido desde el inicio del experimento. La tasa de elongación correspondió a la pendiente de la curva de regresión que relaciona ambas variables (Di Benedetto y Tognetti, 2016).
Intercambio gaseoso: en dos oportunidades durante el periodo experimental se realizaron monitoreos de intercambio gaseoso cada hora, durante un periodo de 24 h, iniciando a las 16:00 h y finalizando a las 11:00 h del día siguiente. El comportamiento diurno y nocturno de intercambio gaseoso se realizó utilizando plantas de edad intermedia, 10-20 años, hojas del estrato medio y la sección intermedia de la hoja. Las lecturas se realizaron con la ayuda de un equipo portátil de intercambio gaseoso marca LiCor6400XT (LiCor Biosciences, Lincoln, NB), utilizando un flujo de aíre de 400 µmol s-1, concentración de entrada de CO de 370 μmol m-2s-1. La actividad fotosintética se de aparición foliar se calculó el filocrono o periodo de tiempo entre la aparición de hojas expresó en μmol de CO2 m-2s-1.
Diseño experimental y análisis estadísticos:El diseño experimental utilizado fue completamente al azar con tres repeticiones, correspondientes a los tres grupos de edad de las plantas. Para los análisis estadísticos se utilizó el paquete estadístico SAS v9.4, realizando un análisis de varianza usando un nivel de significancia del 0,05 y pruebas de separación con la prueba HSD de Tukey. Para el análisis del crecimiento por estratos de la planta, se utilizó un diseño anidado o jerárquico, donde los estratos se consideraron anidados dentro de la edad de las plantas. Se realizaron también análisis de regresión para determinar las tasas de crecimiento foliar, relacionando las variables de crecimiento con el tiempo cronológico transcurrido.
RESULTADOS
Crecimiento foliar
Al iniciar la investigación, las plantas de fique presentaban en promedio 64,89 hojas, y transcurrido el período experimental (154 días), contaban con 83,67 hojas, lo que indica un incremento de 18,78 hojas por planta durante el periodo. El seguimiento al número de hojas permitió calcular la tasa de aparición foliar (TAF) mediante el ajuste de una función lineal entre el número acumulado de hojas emitidas a través del tiempo cronológico, lo cual indicó que bajo las condiciones ambientales del sitio experimental, se registró una TAF promedia de 0,156 hojas día-1, con lo cual la tasa de emisión foliar o filocrono promedio fue de 8,65 días hoja-1, sin variaciones significativas (p<0,05) entre los diferentes grupos de edades de las plantas. En cuanto al crecimiento de las hojas, los análisis realizados indicaron diferencias significativas (p<0,05) entre los estratos o tercios en los cuales se dividieron las plantas, en donde las mayores ganancias en longitud y ancho de las hojas se observaron en el estrato superior de las plantas.
Con lo anterior, las hojas desarrolladas en la parte superior de la planta alcanzaron una longitud final superior (165,9 cm), al alcanzado por las hojas en los estratos medio y bajo de la planta (156,1 y 155,7 cm, respectivamente). Similarmente, el ancho de las hojas, fue superior en los estratos superior y medio de la planta, alcanzando al final del periodo experimental valores de 18,1 y 18,0 cm, que fueron significativamente superiores a los alcanzados en el estrato bajo de la planta (17,2 cm). Por el contrario, en cuanto al grosor de las hojas, no se observaron diferencias en los tres estratos de la planta (p<0,05), finalizando el periodo experimental con valores de 2,44, 2,33 y 2,53 cm para los estratos superior, medio y bajo, respectivamente.
El cálculo de la tasa de elongación foliar (TEF), permitió observar mayores valores promedios en el estrato superior de la planta, que explican la superioridad en el tamaño de las hojas en los estratos superiores (Tabla 1).
| Estrato de la planta | Longitud de la hoja | Ancho de la hoja | Grosor de la hoja |
| Superior | 7,94a | 1,02a | 0,24a |
| Medio | 1,14b | 0,83a | 0,19a |
| Bajo | 0,65b | 0,25a | 0,29a |
| Promedio | 3,31 | 0,70 | 0,24 |
| CV% | 16,88 | 22,91 | 8,64 |
| D. Estándar | 0,29 | 0,24 | 0,07 |
Con los datos de crecimiento foliar del estrato superior se ajustaron modelos de regresión a través de los cuales se muestra una aparente periodicidad de los procesos de crecimiento de las hojas, en donde en primera instancia se alcanzan los valores máximos de longitud, para luego continuar con las ganancias en ancho de las hojas y finalizar con las ganancias en grosor. Los resultados mostraron que los máximos valores de longitud de las hojas se alcanzaron a los 114,48 días, los de ancho foliar a los 127,00 días y los máximos de grosor a los 143,25 días (Tabla 2).
| Variable | Ecuaciones de regresión de mejor ajuste | R2 | Edad (días) a Max. valor |
| Longitud (cm) | Y=34,727+0,8472x-0,0037x2 | 0,9415** | 114,48 |
| Ancho (cm) | Y=1,9292+0,0762x-0,0005x2 | 0,9797** | 127,00 |
| Grosor (cm) | Y=-0,1206+0,0573x-0,0002x2 | 0,8885* | 143,25 |
Intercambio gaseoso
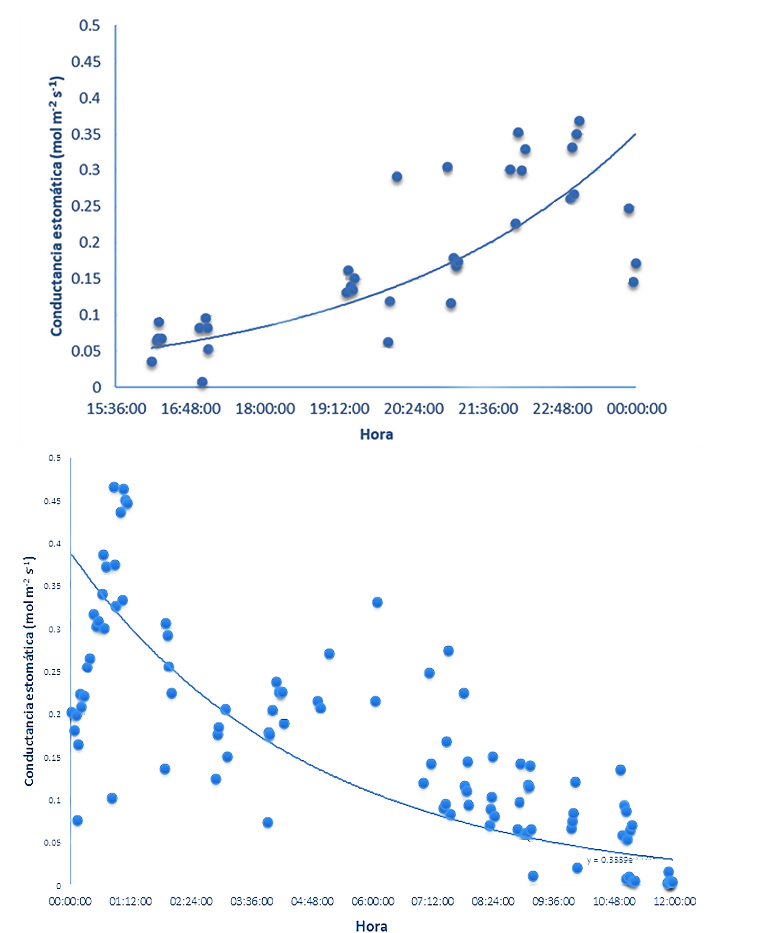
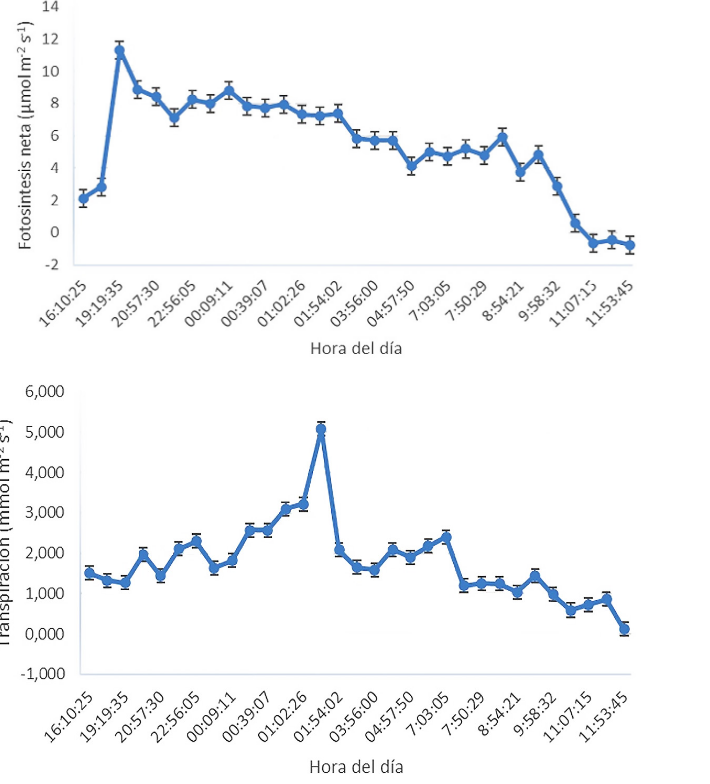
El monitoreo de las tasas de intercambio gaseoso permitió determinar que aun cuando se detectan valores positivos de conductancia estomática (gs) durante ambos periodos, de luz y oscuridad, los resultados mostraron que la mayor apertura se inicia al final del día, durante el periodo de luz, a aproximadamente las 15:30 h. A partir de esa hora, la gs se incrementa sustancialmente y se mantiene durante todo el periodo de oscuridad, para posteriormente decrecer a partir de las 09:30 h, del día siguiente, durante el periodo de luz. El anterior comportamiento sugiere un periodo nocturno de alta conductancia estomática, el cual tiene una duración aproximadamente 18 h, seguido de un período diurno de baja conductancia con una duración aproximada de 6 h (Figura 1).
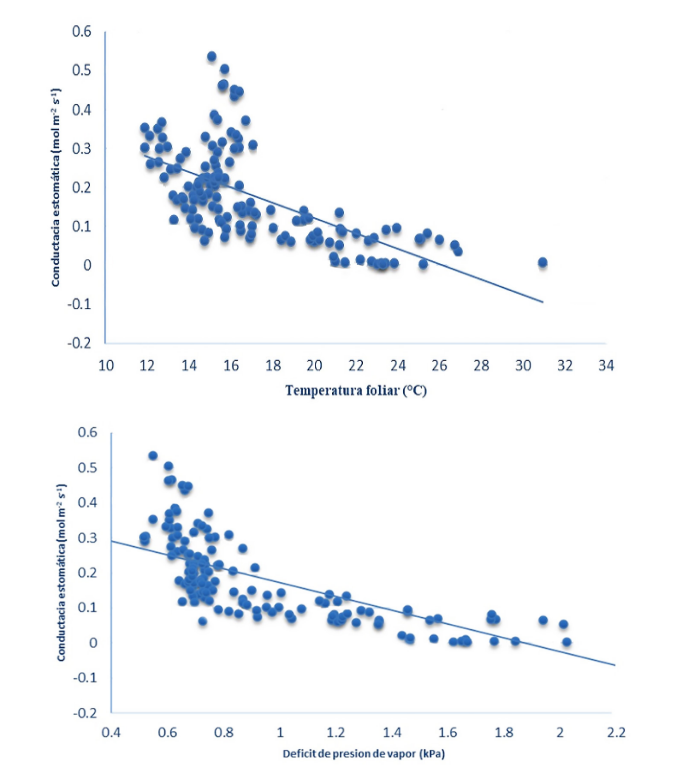
Los anteriores movimientos estomáticos guardaron una estrecha relación con el comportamiento diario del déficit de presión de vapor (VPD) y la temperatura foliar. Se observó que el cierre estomático ocurre cuando la temperatura foliar alcanza valores ≥25,4 °C, y el DPV ≥1,8 kPa (Figura 2).
Lo anterior sugiere que las bajas tasas de conductancia (gs) ocurren por efecto de los incrementos en la temperatura foliar durante las horas de luz, lo cual ocurre probablemente con el fin de evitar pérdidas excesivas de agua por efecto de los incrementos en el DPV. Normalmente en las especies vegetales, los incrementos en la temperatura ambiente ocasionan un incremento en los valores de DPV, ocasionado por el gradiente de humedad entre la atmósfera y el interior de las hojas de las plantas, generando un estrés por déficit de agua, que ocasiona el cierre estomático y limita la capacidad fotosintética (Pereira et al., 2021). A este respecto, una gran cantidad de evidencia sugiere que la conductancia estomática y la transpiración se mantienen hasta un umbral de DPV, a partir del cual se reducen debido a altos valores de DPV, lo que trae como consecuencia una cascada de impactos metabólicos posteriores que incluyen una reducción en las tasas de fotosíntesis y reducción en el crecimiento general de las plantas.
Concomitante con los anteriores resultados, en nuestro experimento las tasas de fijación de CO2 mostraron valores máximos y positivos de asimilación neta (PN), durante todo el período de oscuridad y parte del período de luz. Las tasas de PN guardaron una estrecha relación con la gs (Figura 3). Se observaron valores máximos nocturnos 19:26 h, a una temperatura foliar de entre 15,2 y 19,9 °C. Un comportamiento diario similar fue reportado en Agave por Pimienta-Barrios et al. (2005; 2006), quienes lo asociaron con el comportamiento típico de plantas con metabolismo CAM, que muestran tasas de fotosintesis durante la noche, y de respiración durante el día. Las plantas con metabolismo de PN equivalentes a 12,2-14,5 µmol m-2s-1, CAM presentan apertura estomática durante las cuales se registraron entre las 19:19 y las el periodo de oscuridad a fin de evitar pérdidas de agua por transpiración, fijan en la noche el CO2 atmosférico y lo convierten en ácidos orgánicos, principalmente malato, que es almacenado en las vacuolas (Lüttge, 2010; Andrade et al., 2007). Estos ácidos orgánicos ocasionan una reducción sustancial en el pH al interior de las hojas, acorde a lo reportado en plantas de fique por Casierra-Posada y González, (2009).
Durante el período de luz, los estomas se cierran y los ácidos orgánicos acumulados se movilizan hacia el citoplasma donde se decarboxilan, liberando el CO2 almacenado lo que genera una alta concentración interna, que permite su rápida fijación a través de la vía normal de rubisco, a través del ciclo de Calvin-Benson (Andrade et al., 2007).
Se realizó una correlación entre la PN y las variaciones en la temperatura foliar, lo que indicó que las mayores tasas de fotosíntesis en el cv Ceniza se presentan en la noche a una temperatura foliar de 15 a 18 °C. Las tasas PN se mantienen durante todo el periodo de oscuridad y solamente se reducen al día siguiente, con el aumento en los niveles de luz y la temperatura ambiente. Las tasas de PN se hacen mínimas, durante el periodo de luz, a una temperatura ≥26 °C (Figura 4). El anterior comportamiento corrobora estudios previos que indican que las temperaturas óptimas para la fijación del CO2 en plantas con metabolismo CAM se registra alrededor de los 15 °C. En especies con metabolismo CAM, como Kalanchoë daigremontiana y K. pinnata, se han determinado valores óptimos de la temperatura ambiente que favorecen la acumulación de ácidos orgánicos durante el periodo de oscuridad (Pereira et al., 2021). En varias especies con metabolismo CAM se ha reportado valores óptimos de la temperatura de 25 y 15 °C, en el día y la noche (Yamori et al., 2014)
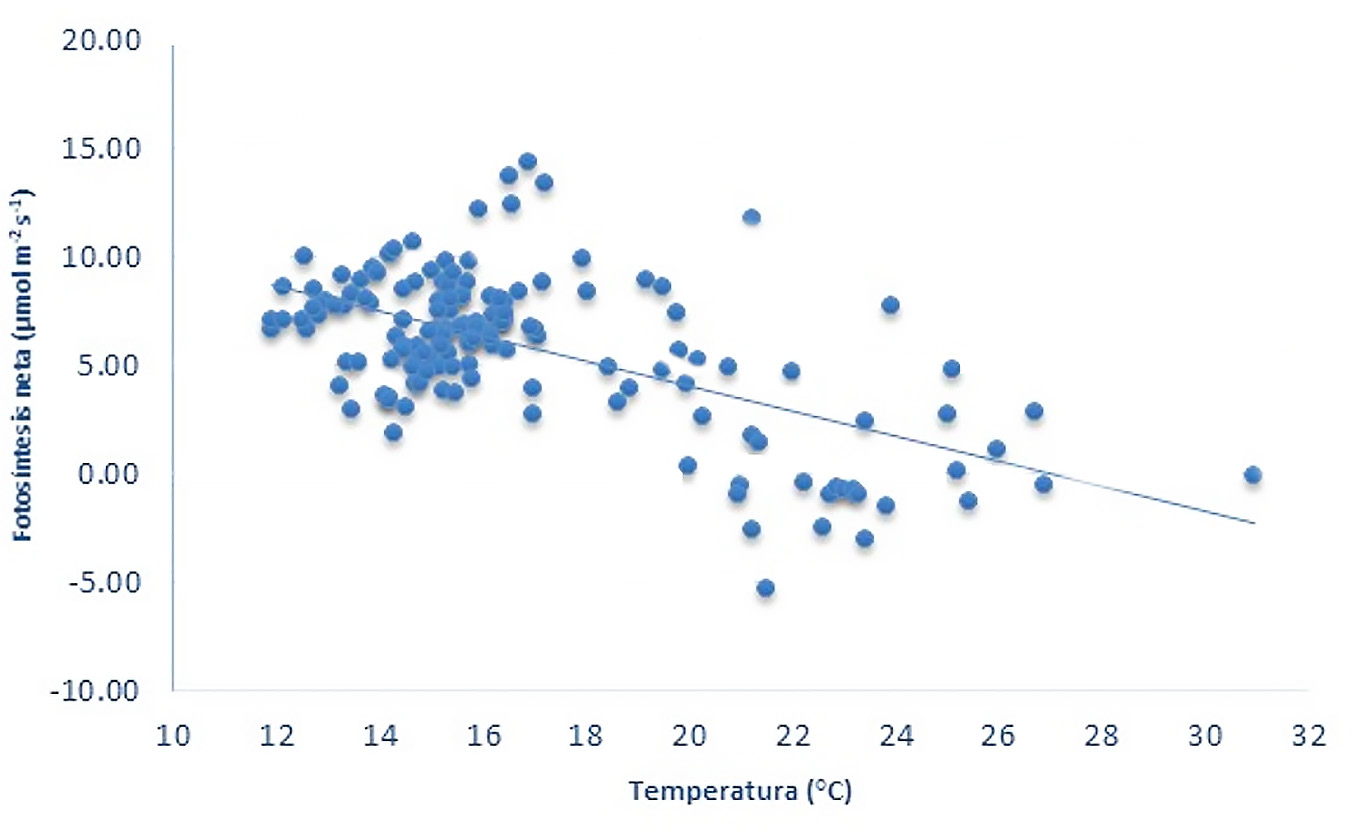
En especies similares al fique, con metabolismo CAM, como es el caso de Agave tequilana (Pimienta-Barrios et al., 2005), se ha demostrado la relación entre la temperatura y las tasas de asimilación de CO2. Estos autores reportan reducciones hasta del 70% a consecuencia de las temperaturas altas durante el día o la noche, lo cual coincide con los resultados obtenidos en este experimento. Lo anterior se explica debido a que en plantas con metabolismo CAM, las temperaturas nocturnas altas ocasionan una reducción en la actividad de la enzima PEP-Carboxilasa (Pimienta-Barrios et al., 2005). Este comportamiento es típico también de plantas que se adaptan al estrés hídrico, las cuales reducen la conductancia estomática durante el período de luz para evitar las perdidas por transpiración debido a las altas temperaturas y alta radiación que ocurren durante el día (Schymanski y Zwieniecki, 2013; Shahzad et al., 2022; Winter y Smith, 2022). El comportamiento diario de la transpiración en nuestros estudios corrobora el comportamiento CAM de las plantas de fique cv Ceniza. Mediante estudios histológicos realizados en el mismo sitio experimental por Murillo-Serna et al. (2018), corroboraron que las plantas de este cultivar presentan características anatómicas de plantas adaptadas a la sequía y metabolismo CAM. Similares resultados de anatomía foliar y distribución de los cloroplastos fueron obtenidos para plantas tipo CAM por Olmedilla Arnal et al. (2010).
CONCLUSIONES
Nuestros resultados permiten concluir que las plantas de fique del cultivar ceniza presentan una zona de activo crecimiento que se ubica en el estrato superior de la planta, en donde ocurre una activa emisión de nuevas hojas, las cuales registran constantes ganancias en longitud y ancho de las hojas. Las hojas desarrolladas en el estrato superior de la planta presentaron tasas superiores de crecimiento, con lo cual alcanzaron una mayor longitud y ancho de las hojas. Las plantas de fique cv Ceniza presentaron un comportamiento típico de metabolismo CAM, en donde las tasas máximas de captura de CO2 se registraron durante el periodo de oscuridad, y se mantienen durante aproximadamente 18 h. Las tasas de intercambio gaseoso parecen responder al comportamiento diario de la temperatura foliar y el déficit en la presión de vapor. Las mayores tasas de fotosíntesis se registran a temperaturas foliares entre 15 y 18 ºC.
Agradecimientos
Agradecimientos
Este trabajo formó parte del macroproyecto “Manejo Integrado e industrialización del cultivo de Fique”, producto “Caracterización del desarrollo foliar de los principales cultivares de fique y su relación con el ambiente,” desarrollado en ejecución del Convenio 1810/13 suscrito entre el Ministerio de Agricultura y Desarrollo Rural y la Corporación Colombiana de Investigación Agropecuaria- AGROSAVIA. Los autores gradecen al estudiante de la Universidad de Antioquia Jhon Steven Murillo Serna quien como parte de su trabajo de pasantía realizó la toma de datos en campo.
REFERENCIAS
Agronet, 2020. Área sembrada y área cosechada del cultivo de fique en Colombia. https://www.agronet.gov.co/estadistica/ Paginas/home.aspx?cod=1
Anandjiwala, R. D. and John, M. 2010. Sisal- cultivation, processing, and products. Industrial Applications of Natural Fibres: Structure, Properties and Technical Applications, 181-95.
Andrade, J. L., de la Barrera, E., Reyes Garcia, C., Ricalde, M. F., Vargas Soto, G. y Cervera, J. C. 2007. El metabolismo acido de las crasulaceae: diversidad, fisiología ambiental y productividad. Boletín de la Sociedad Botánica de México, 81, 37–50.
Casierra-posada, F. y Gómez, N. E. 2008. Crecimiento foliar y radical en plantas de fique (Furcraea castilla . F. macrophylla) bajo estrés por encharcamiento. Agronomia Colombiana, 26(3):381–388.
Casierra-Posada, F. y González, D. M. 2009. Cambio circadiano de pH y acidez titulable en la savia de fique (Furcraea castilla . F. macrophylla). Orinoquia, 13(1): 5–13.
Di Benedetto, A. y Tognetti, J. 2016. Técnicas de análisis de crecimiento de plantas: su aplicación a cultivos intensivos. Ria, 42(3): 258–282. http://www.scielo.org.ar/pdf/ria/v42n3/v42n3a07.pdf
Eggli, U. and Nyffeler, R. 2020. Monocotyledons. In Monocotyledons: Illustrated Handbook of Succulent Plants. Springer. https://doi.org/10.1007/978-3-662-56486-8
Lüttge, U. 2010. Ability of crassulacean acid metabolism plants to overcome interacting stresses in tropical environments. AoB PLANTS, 2010: 1–15. https://doi.org/10.1093/aobpla/plq005
Murillo-Serna, J.S., Rincón-Baron, E.J. y Alzate-Guarín, F. 2018. Anatomía foliar comparativa de tres especies de Furcraea (Asparagaceae : Agavoideae). Hoehnea, 45(4): 607–615.
Olmedilla Arnal, A., Alché Ramírez, J. D. D. y Rodríguez García, M. I. 2010. Identificación histológica y ultraestructural de plantas C 4 y CAM. C4 y CAM: Características Generales y Uso en Programas de Desarrollo de Tierras Áridas y Semiáridas. 107–114.
Pereira, P. N., Niechayev, N. A., Blair, B.B. and Cushman, J. C. 2021. Chapter 10 Climate Change Responses and Adaptations in Crassulacean Acid Metabolism (CAM) Plants. https://doi.org/10.1007/978-3-030-64926-5_10
Pimienta-Barrios, E., Zañudo-Hernández, J. and García-Galindo, J. 2006. Fotosíntesis estacional en plantas jóvenes de Agave tequilana. Agrociencia. 40(6): 699–709.
Pimienta-barrios, E., Zañudo-Hernández, J., Nobel, P. S. y García-Galindo, J. 2005. Respuesta fisiológica a factores ambientales del agave azul (Agave. In scientiaCUCBA. 7(2).
Shahzad, S., Hussain, M., Arfan, M. and Munir, H. 2022. Physiological and biochemical attributes of agave sisalana resilient adaptation to climatic and spatio-temporal conditions. Pakistan Journal of Botany. 54(1): 169–178. https://doi.org/10.30848/PJB2022-1(15)
Winter, K. and Smith, J. A. C. 2022. CAM photosynthesis: the acid test. New Phytologist, 233(2): 599–609. https://doi.org/10.1111/nph.17790
Schymanski, S.J. and Maciej Zwieniecki, O.D. 2013. Stomatal Control and Leaf Thermal and Hydraulic Capacitances under Rapid Environmental Fluctuations. PLoS ONE 8(1): e54231. https://doi.org/10.1371/journal.pone.0054231
Yamori, W., Hikosaka, K. and Way, D. A. 2014. Temperature response of photosynthesis in C3, C4, and CAM plants: temperature acclimation and temperature adaptation. Photosynthesis Research, 119(1): 101-117.
Yu, C. 2005. Sisal. In Bast and other plant fibres. Woodhead Publishing. 228-273.

